All published articles of this journal are available on ScienceDirect.

Entomopathogenic Efficacy and Cuticle-degrading Enzyme Production in Beauvaria bassiana and Verticillium lecanii Isolates Targeting Adult Cowpea Weevils
Authors Info & Affiliations
Abstract
Introduction
Cowpea weevils (Callosobruchus maculatus) are a major post-harvest pest responsible for significant quantitative and qualitative losses in stored cowpea across tropical regions. The growing concerns associated with synthetic insecticides, including environmental contamination, pest resistance, and human health risks, have intensified the search for sustainable biological alternatives. This study evaluated the efficacy of two entomopathogenic fungi, Beauveria bassiana and Verticillium lecanii, against adult cowpea weevils and examined the role of their cuticle-degrading enzymes in pest control.
Methods
Contact bioassays were conducted to assess adult weevil mortality and suppression of first-generation (F1) emergence following treatments with fungal isolates. Standard enzymatic assays were performed to determine the specific activities of protease, lipase, and exochitinase produced by the fungi isolates.
Results
Both fungi isolates induced progressive mortality in cowpea weevils, with B. bassiana demonstrating comparatively higher pesticidal activity. In addition, treatments with both fungi isolates significantly reduced F1 adult emergence, with emergence levels remaining below the 50% threshold considered indicative of effective population suppression. Enzymatic analysis showed that both fungi isolates produced measurable cuticle-degrading enzymes. V. lecanii exhibited relatively higher protease and lipase specific activities, whereas B. bassiana showed higher exochitinase activity.
Discussion
The observed pesticidal effects are attributable to the production of cuticle-degrading enzymes, which facilitate fungal penetration and infection of the pest host. Differences in enzyme activity between the isolates may explain variations in their pathogenicity.
Conclusion
Beauveria bassiana and Verticillium lecanii showed potential as environmentally sustainable biocontrol agents for managing cowpea weevil infestations in stored cowpea.
1. INTRODUCTION
Cowpea (Vigna unguiculata (L.) Walp.) remains one of the most economically and nutritionally significant grain legumes cultivated in sub-Saharan Africa, where it serves as a primary source of dietary protein, essential amino acids, vitamins, and minerals for millions of people. Beyond its nutritional contribution, cowpea supports soil fertility through biological nitrogen fixation and contributes substantially to household income and livestock feed systems [1]. Countries such as Nigeria, Niger, and Burkina Faso dominate global production, with Nigeria leading global production per year at approximately 4.3 million tons from the global total of 9.8 million tons, accounting for 61% of Africa’s output and 48% worldwide [2]. Despite its agronomic and socio-economic importance, cowpea production and storage are severely constrained by insect pests, particularly the cowpea weevil, Callosobruchus maculatus, which is recognized as the most destructive post-harvest pest of cowpea in tropical regions, causing losses of up to 60% or more during storage [3].
Infestation by cowpea weevils begins in the field but escalates during storage, where rapid multiplication under favorable temperature and humidity conditions may result in 50-100% grain loss within a few months if untreated [4]. Adult females oviposit on the seed surface; upon hatching, larvae bore into the cotyledons, completing development with the grain and emerging as adults, thereby reducing seed weight, market value, germination potential, and nutritional quality. In heavily infested stocks, contamination with insect fragments and frass further compromises food safety and consumer acceptability [5]. Given that cowpea is predominantly produced by smallholder farmers who rely on traditional storage systems, the management of storage pests remains a critical food security challenge across West Africa.
For decades, the primary strategy for controlling storage pests has relied on synthetically chemical insecticides, including organophosphates and pyrethroids. While these compounds provide rapid knock-down and broad-spectrum activity, their indiscriminate and repeated use has raised serious concerns regarding pesticide residue, environmental contamination, pest resistance, and human health risks [6]. Several toxicological studies have linked chronic pesticide exposure to oxidative stress, endocrine disruption, neurodegenerative disorders, and carcinogenesis [7, 8]. In addition, insect populations of storage pests have demonstrated varying degrees of resistance to conventional insecticides, thereby reducing long-term efficacy and necessitating higher application rates [9]. These limitations underscore the urgent need for sustainable, eco-friendly, and target-specific alternatives for managing cowpea weevil.
Biological control using microbial agents has emerged as a promising component of Integrated Pest Management (IPM) strategies. Among microbial biopesticides, entomopathogenic fungi have attracted considerable attention because of their broad host range, environmental compatibility, and unique infection of the host. Notably, Beauveria bassiana and Verticillium lecanii have been widely studied for their pathogenicity against lepidoptera insects [10]. However, there is little literature on the pathogenicity of these fungi in the coleopteran order, such as the cowpea weevil. These fungi are naturally occurring soilborne microorganisms capable of infecting insects through direct contact with the cuticle, thereby making them particularly suitable for controlling external and storage pests [11].
A critical determinant of fungal virulence is the secretion of cuticle-degrading enzymes, including proteases, chitinases, and lipases, which collectively disrupt the integrity of the insect integument. The insect cuticle is primarily composed of chitin fibres embedded in a protein-lipid matrix; thus, enzymatic hydrolysis of macromolecules facilitates fungal entry and colonization [12].
Although numerous studies have evaluated the insecticidal potential of B. bassiana and V. lecanii against field pests such as aphids and whiteflies, comparatively fewer investigations have focused specifically on adult cowpea weevils under storage. Therefore, given the pressing need for safer alternatives to chemical insecticides and the pivotal role of cuticle-degrading enzymes in fungal pathogenicity, this study investigated the efficacy of B. bassiana and V. lecanii against adult cowpea weevils infesting stored cowpea.
2. MATERIALS AND METHODS
2.1. Collection of the Fungal Isolates
B. bassiana and V. lecanii isolates used in this study were obtained from the Microbial Reference Laboratory Unit of Biocrops Biotechnology Limited, Abuja, Nigeria. The isolates were originally recovered from soil samples collected within the Abuja environment and had previously been identified and confirmed through molecular characterization. For culturing, fungal isolates were inoculated onto Potato Dextrose Agar (PDA) supplemented with chloramphenicol to suppress bacterial contamination. The inoculated plates were incubated at 28 ± 2°C for 72 hours under ambient laboratory conditions. After incubation, distinct colonies that developed on the agar surface were discretely isolated and repeatedly sub-cultured on freshly prepared PDA plates to obtain pure cultures. The purified isolates were further confirmed through morphological observation and biochemical characterization. Confirmed isolates were maintained on agar slants and stored at 4°C. Subsequently, the isolates were subjected to liquid fermentation to enhance microbial proliferation, using sucrose solution as a carbon-rich substrate and liquid growth medium.
2.2. Collection of Cowpea Seeds and Cowpea Weevils
The black-eyed pea commercialized variety (Nigeria Sampea-10) cowpea seeds were obtained directly from the experimental farm. The seeds were harvested directly from the field to minimize the likelihood of prior exposure to pesticides or storage chemicals. Immediately after collection, the seeds were aseptically packaged in sterile bags and placed in tightly sealed containers to prevent contamination. The samples were subsequently stored under freezing conditions until required for experimental use.
To obtain the cowpea weevils, cowpea seeds naturally infested with cowpea weevils were collected from storage facilities and local traders within Abuja. Adult weevils of both sexes emerging from visibly infested seeds were isolated through selective hand-picking and transferred to clean containers for subsequent experimental procedures.
2.3. Rearing of Cowpea Weevils
Adult cowpea weevils obtained from naturally infested cowpea seeds were cultured under ambient laboratory conditions to establish a stable insect colony for experimental use. The rearing procedure followed the method described by Nwosu et al. [13]. The newly emerged first filial generation (F1) of cowpea weevils (male and female) at the ratio 50:50 was collected and used for all subsequent experimental bioassays. Insect handling complied with institutional guidelines and standards.
2.4. Evaluation of the Insecticidal Activity of the Fungal Isolates Against Cowpea Weevil
The microbial load of B. bassiana and V. lecanii isolates was determined using a ten-fold serial dilution technique followed by total viable plate count, as previously described [14]. Colony-Forming Units (CFU) were enumerated to estimate the concentration of fungal suspension used in subsequent bioassays. The insecticidal activity of B. bassiana and V. lecanii isolates against cowpea weevils was evaluated using the contact method with all treatments conducted in triplicate. Approximately 20 grams of cowpea seeds were introduced into sterile transparent plastic 0.5 L containers fitted with perforated lids to allow adequate ventilation. Ten adult cowpea weevils were then introduced into each container. Subsequently, the seeds and insects were treated by spraying 1-4 mL of liquid formulation of the fungal isolates at concentrations of 2.56 x 104 CFU/ml and 4.4 x 106 CFU/ml of B. bassiana and V. lecanii, respectively. Spraying was done once using sterile 10 mL plastic spray bottles to ensure uniform distribution of the fungal suspension over seeds and insects. Following treatment, the containers were maintained under ambient laboratory conditions and monitored for insect mortality during the experimental period.
Adult mortality of cowpea weevils was monitored at 24-hour intervals throughout the exposure period. Observations were conducted for 12 days post-treatment. At each observation period, dead weevils were carefully removed from the containers using forceps, with the aid of a magnifying lens when necessary to detect individual weevils concealed among the cowpea seeds. Adult cowpea weevils that showed no movement or response to gentle probing with forceps were considered dead and were subsequently removed from the experimental units to prevent recounting. The percentage mortality of adult cowpea weevils was determined using the formula in Eq. (1):
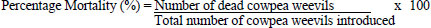 ...(1)
...(1)
This procedure was repeated for all treatments and replicated throughout the experimental period.
2.5. Evaluation of the Suppressive Effect of the Fungal Isolates Treatment on Cowpea Weevil Population Development
The suppressive effect of the fungal isolates treatments on cowpea weevil population development was determined by assessing the emergence of first-generation (F1) adult cowpea weevils, forty (40) days after treatment, using the same experimental units employed for the mortality assessment. At the end of the daily observation period for 40 days, newly emerged adult weevils were counted and recorded for each treatment. The percentage emergence (PE) of F1 adults was calculated using the formula in Eq. (2):
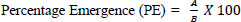 ...(2)
...(2)
Where A = Number of adult insects that emerged in treated samples, B= Number of adult insects that emerged in the control treatment.
Treatments that resulted in ≤50% adult emergence relative to the control were considered to exhibit promising inhibitory effects on the development of the pest population [15].
2.6. Assay of the Activity of the Cuticle Degrading Enzymes
2.6.1. Extracellular Enzyme Production of the Fungal Isolates
The nutrient broth was prepared according to the manufacturer’s (HiMedia) specifications. For the production of the extracellular enzymes as reported previously [16], 1ml of the liquid medium of the isolates containing 2.56 x 104 CFU/ml for Beauveria bassiana and 4.4 x 106 CFU/ml for Verticillium lecanii was incubated in the nutrient broth at room temperature (28 ± 2°C) for 96 hours. After incubation, the contents of each test tube were centrifuged at 10,000 rpm for 20 minutes at 4°C. Finally, the supernatants were used in enzymatic assays.
2.6.2. Total Protease Assay
Total protease activity was assayed using the substrate casein. The reaction mixture consisted of 100 µL of Tris-HCl buffer (20 mM, pH 8), 30 µL of the substrate (20 mg/mL), and 30 µL of enzyme solution. After thoroughly mixing and incubating at 37°C for 100 minutes, the reaction was stopped by adding 100 µL of trichloroacetic acid (30% w/v), then cooling to 4°C for 45 minutes. The unhydrolyzed substrate was precipitated by centrifugation at 10000rpm for 10 minutes at 4°C. Finally, the Folin-Ciocalteu reagent (50 µL) was added to the supernatant, and absorbance was measured at 630 nm using a UV/VIS spectrophotometer (1800-series, Shimadzu). Tyrosine solutions were used as a standard curve for enzyme activity calculation.
2.6.3. Lipase Assay
Lipase activity was assayed using p-nitrophenyl butyrate as substrate. For the assay, 100 µL of Tris-HCl buffer (20 mM, pH 7.5), 50 µL of p-nitrophenyl butyrate, and 30 µL of enzyme solution were thoroughly mixed at 25°C for 5 minutes. Then, 100 µL of sodium hydroxide (1 N) was added to the reaction mixture. Finally, absorbance was read at 405 nm by a UV/VIS spectrophotometer (1800-series, Shimadzu). Calculation of enzyme activity was performed using the calibration curve of p-nitrophenol solutions, as a reaction product.
2.6.4. Exochitinase Assay
The exochitinase activity was assayed using p-nitrophenyl-N-acetyl-β-D-glucosaminide as substrate. For the assay, 30 µL of phosphate buffer (50 mM, pH 4.8), 30 µL of p-nitrophenyl-N-acetyl-β-D-glucosaminide, and 30 µL of enzyme solution were mixed and incubated for 10 minutes at 37°C. The reaction was stopped by adding sodium carbonate (30 µL, 0.4 M), and the absorbance was recorded at 400 nm on a UV/VIS spectrophotometer (1800-series, Shimadzu). The enzyme activity was calculated based on the extinction coefficient equal to 18.5 mM− 1. cm− 1.
2.6.5. Protein Concentration Assay
The total soluble protein concentration in the microbial isolate samples was determined using the previously described method [17] with bovine serum albumin (BSA 1 mg/ml) as the standard.
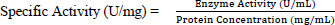 ...(3)
...(3)2.7. Statistical Analysis
Data were expressed as mean value ± standard error of the mean (S.E.M) and analyzed using ANOVA. Significant differences between the control and treatment groups, as well as differences in enzyme activities between fungal groups, were determined using Duncan’s Multiple Range Test (DMRT) in SPSS (version 26). Mean values with different superscript letters were considered significantly different at p < 0.05.
3. RESULTS
The efficacy study revealed the mortality potential of the liquid formulation of the two fungal isolates on cowpea weevil. The results of the efficacy of Beauvaria bassiana and Verticillium lecanii on the mortality of adult cowpea weevils are shown in Figs. (1 and 2), respectively. The findings indicate that the fungal isolates induced varying degrees of mortality in the cowpea weevils, with effectiveness increasing over time and at higher microbial concentrations. B. bassiana demonstrated the highest mortality rate. Comparisons were made among treatment concentrations at each exposure time and across exposure times within the same treatment concentration.
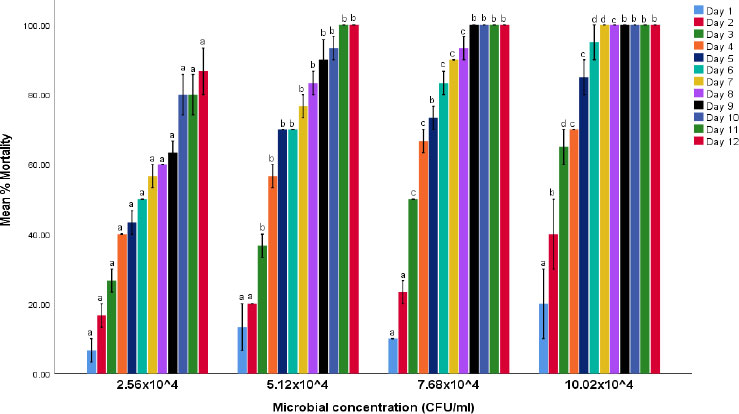
Percentage mortality of cowpea weevils treated with liquid formulation of Beauvaria bassiana isolate.
Note: Values are mean percentage mortality ± S.E.M. S.E.M = Standard error of the mean. Values between experimental treatments in each microbial concentration bearing the same superscript are not statistically different at the 5% level (P>0.05).
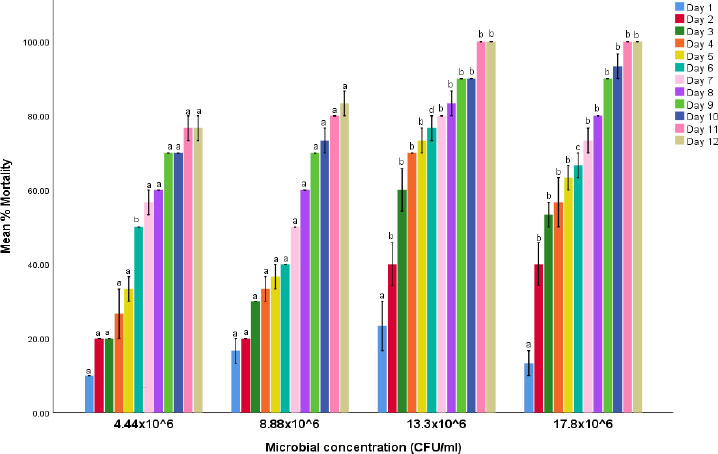
Percentage mortality of cowpea weevils treated with liquid formulation of verticillium lecanii isolate.
Note: Values are mean percentage mortality ± S.E.M. S.E.M = Standard error of mean. Values between experimental treatments in each microbial concentration bearing the same superscript are not statistically different at the 5% level (P>0.05).
The percent (%) emergence of the first generation of the adult cowpea weevil after forty days of assessing the percent mortality of the cowpea weevils was revealed. Treatments with first-generation emergence ≤ 50% are considered promising [15]. Tables 1 and 2 show the percentage emergence of the first generation (F1) of cowpea weevil treated with the liquid formulation of the fungal isolates, respectively, demonstrating the promising potential of each concentration used in the study. The findings indicate significant reduction in F1 emergence across isolate treatments.
| Fungal Isolate | Treatment Dose (ml) | TreatmentConcentration (CFU/ml) | Mean Emergence of Treatment (X) | Mean Emergence of Control (Y) | PE(%) | Remarks |
|---|---|---|---|---|---|---|
| Beauveria bassiana | 1 | 2.56x104 | 5.01±0.01 | 11.3±0.04 | 44.33 | Promising |
| 2 | 5.12x104 | 4.00±0.02 | 9.66±0.11 | 41.40 | Promising | |
| 3 | 7.68x104 | 3.00±0.01 | 7.67±0.05 | 39.11 | Promising | |
| 4 | 10.2x105 | 2.33±0.11 | 6.33±0.03 | 36.80 | Promising |
Note: Values represent the mean number of newly emerged F1 adult cowpea weevils ± S.E.M. S.E.M= Standard error of the mean. Where X = Mean number of newly emerged adult insects in the treatment samples; and Y =Mean number of newly emerged adult insects in the untreated control samples. Treatments where first-generation emergence is ≤ 50% are considered promising. PE= Percentage emergence.
| Fungal Isolate | Treatment Dose (ml) | TreatmentConcentration (CFU/ml) | Mean Emergence of Treatment (X) | Mean Emergence of Control (Y) | PE(%) | Remarks |
|---|---|---|---|---|---|---|
| Verticillium lecanii | 1 | 4.44x106 | 3.00±0.02 | 11.3±0.04 | 26.54 | Promising |
| 2 | 8.88x106 | 2.00±0.05 | 9.66±0.11 | 20.70 | Promising | |
| 3 | 13.3x106 | 1.00±0.01 | 7.67±0.05 | 13.03 | Promising | |
| 4 | 17.8x106 | 0.33±0.01 | 6.33±0.03 | 5.21 | Promising |
Note: Values represent the mean number of newly emerged F1 adult cowpea weevils ± S.E.M. S.E.M = Standard error of mean. Where X = Mean number of newly emerged adult insects in the treatment samples; and Y =Mean number of newly emerged adult insects in the untreated control samples. Treatments where first-generation emergence is ≤ 50% are considered promising. PE= Percentage emergence.
The assessment of the cuticle-degrading enzyme activity of the fungal isolates revealed protease, lipase, and exochitinase activity, suggesting the involvement of these enzymes in the toxicity potential of the fungal isolates toward cowpea weevils. Figures 3, 4, and 5 show the Protease, Lipase, and Exochtinase specific activities of the fungal isolates, respectively. The findings revealed that the fungal isolates exhibited measurable protease, lipase, and exochitinase activity at varying levels, with V.lecanii displaying relatively higher protease and lipase-specific activity while B. bassiana showed a significantly higher exochitinase-specific activity.
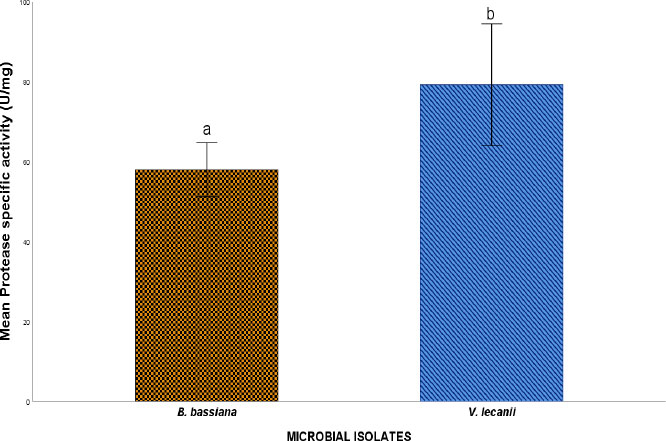
Protease specific activity of microbial isolates.
Note: Values are mean specific enzyme activity ± S.E.M. S.E.M = Standard error of the mean. Values between experimental microbial isolates bearing different superscripts are significantly different at the 5% level (P<0.05).
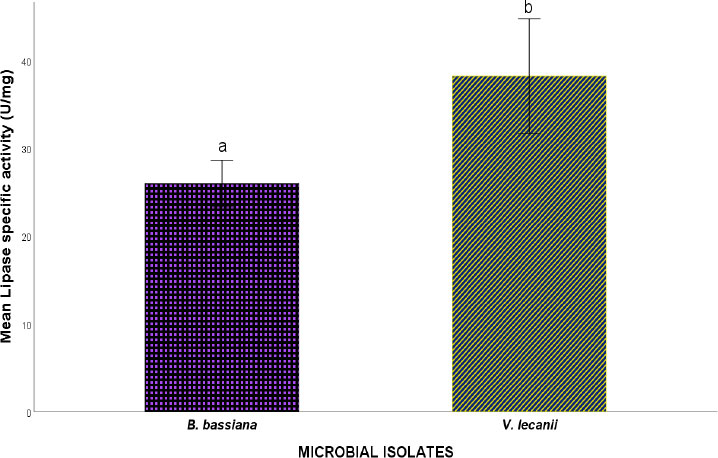
Lipase specific activity of microbial isolates.
Note: Values are mean specific enzyme activity ± S.E.M. S.E.M = Standard error of mean. Values between experimental microbial isolates bearing different superscripts are significantly different at the 5% level (P<0.05).
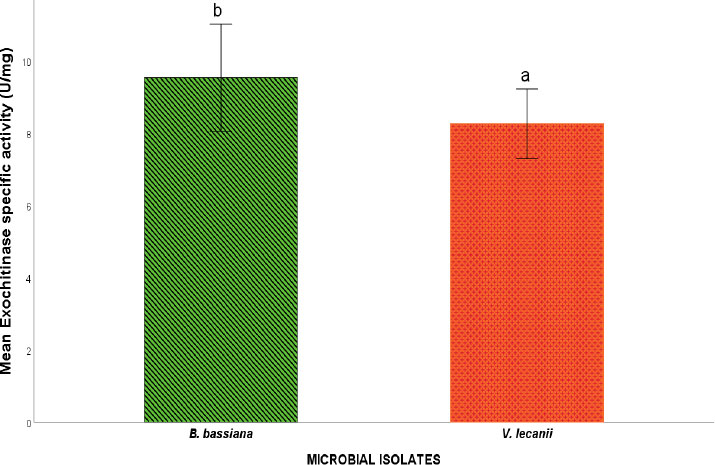
Exochitinase specific activity of microbial isolates.
Note: Values are mean specific enzyme activity ± S.E.M. S.E.M = Standard error of mean. Values between experimental microbial isolates bearing different superscripts are significantly different at the 5% level (P<0.05).
4. DISCUSSION
The present study evaluated the efficacy of the entomopathogenic fungi, Beauveria bassiana and Verticillium lecanii, against adult cowpea weevil (Callosobruchus maculatus) and examined the contribution of their cuticle-degrading enzymes to fungal pathogenicity. The results demonstrate that both fungal isolates exerted significant insecticidal effects, as evidenced by progressive increases in adult mortality, marked suppression of F1 progeny emergence, and measurable activity of key extracellular enzymes involved in insect cuticle degradation. These findings reinforce the growing body of evidence supporting entomopathogenic fungi like B. bassiana and V. lecanii as viable biological control agents for stored-product pests.
The mortality patterns observed in the present study and shown in Figs. (1 and 2) indicate that fungal efficacy increased with exposure duration and microbial concentration, with B. bassiana producing relatively higher adult mortality compared with V. lecanii. This progressive mortality trend is consistent with the typical infection dynamics of entomopathogenic fungi, which require time for conidial adhesion, germination, cuticular penetration, and systemic colonization of the host insect [18]. Unlike chemical insecticides, which often produce rapid knockdown effects, fungal pathogens act through a multi-stage infection process involving mechanical penetration and enzymatic degradation of the insect cuticle followed by proliferation within the haemocoel [19]. Consequently, mortality typically increases gradually over several days, as reflected in the present findings.
The higher mortality induced by B. bassiana may be attributed to its well-documented virulence and adaptability to diverse insect hosts. Several studies have reported strong pathogenic activity of B. bassiana against lepidopteran field pests and a few coleopteran pests, including Sitophilus zeamais and Tribolium castaneum [20, 21]. The relatively lower mortality observed with V. lecanii in comparison to B. bassiana may reflect differences in virulence among fungal species or isolate-specific pathogenicity. Although V. lecanii (also known as Lecanicillium lecanii) is widely recognized for its effectiveness against hemipteran pests such as aphids and whiteflies, its pathogenicity toward coleopteran pests has been reported to vary depending on strain, environmental conditions, and host susceptibility [22, 23].
Beyond direct mortality, an important indicator of pest management efficacy in stored-product systems is the suppression of insect reproduction and population development. In this study, treatments with both fungal isolates significantly reduced the emergence of the first filial generation (F1) of cowpea weevils, with all treatment concentrations producing emergence rates below the 50% threshold considered indicative of promising insecticidal activity. The suppression of F1 progeny suggests that fungal infection not only affects adult survival but may also impair oviposition, egg viability, or larval development within the host seed. Similar reductions in progeny emergence following fungal treatment have been reported in other storage pest systems, where infection of adults leads to physiological stress and reduced reproductive capacity [20]. In addition, fungal growth on infected insects may contaminate oviposition sites, thereby exposing eggs and newly emerged larvae to infectious propagules [24]. Such population-level suppression is particularly advantageous for integrated pest management strategies because it limits pest resurgence during storage.
The pathogenic success of entomopathogenic fungi is largely dependent on their ability to breach the protective insect cuticle, which is composed of chitin microfibrils embedded in a matrix of proteins and lipids [25]. In this study, both B. bassiana and V. lecanii exhibited measurable activities of protease, lipase, and exochitinase enzymes, as shown in Figures 3, 4, and 5, respectively, indicating their capacity to enzymatically degrade the structural components of the insect integument. The secretion of such cuticle-degrading enzymes is widely recognized as a key virulence factor in fungal infection of insects [26].
Interestingly, the enzyme activity profiles observed in this study revealed species-specific differences in enzymatic expression. V. lecanii exhibited relatively higher specific activities of protease and lipase, whereas B. bassiana displayed significantly higher exochitinase activity. Proteases are essential for degrading the proteinaceous components of the insect epicuticle and endocuticle, thereby facilitating fungal penetration into the host body [12]. Lipases, on the other hand, hydrolyze lipid layers present in the insect epicuticle, which represent the first barrier to fungal infection [27]. The higher protease and lipase activity observed in V. lecanii suggests a strong capacity for disrupting the outer cuticular barrier, potentially enhancing the initial stages of fungal attachment and penetration.
Conversely, the elevated exochitinase activity recorded for B. bassiana indicates an enhanced ability to degrade chitin, which forms the structural backbone of the insect cuticle. Chitinolytic enzymes play a critical role during later stages of cuticle penetration by breaking down chitin polymers within the procuticle, thereby enabling fungal hyphae to invade the haemocoel [27, 28]. Previous studies have similarly reported high chitinase activity in virulent strains of B. bassiana, linking this enzymatic capability with enhanced insect pathogenicity [29]. The comparatively higher exochitinase activity observed in B. bassiana in this study may therefore partly explain the greater mortality rates induced by this isolate.
The combined action of proteases, lipases, and chitinases is widely recognized to act synergistically during fungal infection. These enzymes collectively degrade the complex cuticular matrix, allowing fungal germ tubes to penetrate the host integument and establish infection [12]. The presence of all three enzyme types in both fungal isolates in this study therefore supports their functional role in the observed insecticidal activity against cowpea weevils.
From an applied perspective, the demonstrated efficacy of B. bassiana and V. lecanii highlights their potential as eco-friendly alternatives to synthetic insecticides for managing cowpea weevil infestations in stored grains. Biological control agents based on entomopathogenic fungi offer several advantages, including reduced environmental toxicity, lower risk of pesticide residues in food products, and compatibility with integrated pest management strategies [30]. Furthermore, because fungal pathogens infect insects through contact rather than ingestion [23], they are particularly suited for controlling pests that develop within stored grains where conventional insecticides may have limited penetration.
Nevertheless, the practical application of fungal biopesticides in storage systems may be influenced by environmental factors such as temperature, humidity, and formulation stability. Previous studies have shown that fungal virulence and spore viability can be affected by unfavorable storage conditions, highlighting the need for optimized formulations and delivery systems to ensure consistent field performance [11, 24]. Future studies should therefore explore formulation improvements, evaluate the persistence of fungal propagules under storage conditions, and investigate potential synergistic interactions with other biological or botanical control agents.
Overall, the findings of this study demonstrated that B. bassiana and V. lecanii possess significant insecticidal potential against adult cowpea weevils and effectively suppress pest population development through both direct pathogenicity and enzymatic degradation of the insect cuticle. The differential expression of cuticle-degrading enzymes between the two fungal species further highlights the complexity of fungal virulence mechanisms and underscores the importance of enzyme-mediated cuticle penetration in entomopathogenic fungal infection.
5. STUDY LIMITATIONS
While the study demonstrates promising biocontrol potential, the limitation is that the experiments were conducted under laboratory conditions, which may not fully reflect the complexity of a real storage environment, where temperature, humidity, and microbial interactions may vary significantly. Future studies should incorporate field trials and further molecular approaches to elucidate mechanisms and enhance applicability.
CONCLUSION
The study demonstrates that the entomopathogenic fungi Beauveria bassiana and Verticillium lecanii possess significant biocontrol potential against cowpea weevil, as evidenced by increased insect mortality and substantial suppression of F1 adult emergence. The pathogenicity of the isolates appears to be associated with the production of cuticle-degrading enzymes, including proteases, lipases, and exochitinases, which facilitate fungal penetration and infection of the insect host. These findings highlight the potential of these entomopathogenic fungi as environmentally sustainable alternatives to synthetic insecticides for the management of cowpea weevil in stored cowpea. Further studies focusing on formulation optimization and field-scale validation are recommended to support their integration into post-harvest pest management strategies.
AUTHORS’ CONTRIBUTIONS
The authors confirm contribution to the paper as follows: O.K.N., E.O.O., I.K.O. and H.B.: Study conception and design; O.K.N. and E.O.O.: Data collection; D.O.O and O.K.N.: Data Analysis and interpretation of results; O.K.N. and E.O.O.: Draft manuscript; O.K.N., E.O.O., I.K.O., H.B.: Writing, reviewing, and editing. All authors reviewed the results and approved the final version of the manuscript.
LIST OF ABBREVIATIONS
| ANOVA | = Analysis of Variance |
| BSA | = Bovine Serum Albumin |
| CFU | = Colony Forming Unit |
| DMRT | = Duncan’s Multiple Range Test |
| F1 | = First Generation |
| IPM | = Integrated Pest Management |
| PDA | = Potato Dextrose Agar |
| PE | = Percentage Emergence |
| SPSS | = Statistical Package for Social Sciences |
| UV/VIS | = Ultraviolet-Visible Spectroscopy |
AVAILABILITY OF DATA AND MATERIALS
The data supporting the findings of this article were originally generated from the research study presented in this article. The datasets are only available within the authors' and institutional archives.
ACKNOWLEDGEMENTS
The authors appreciate Bio-crops Nigeria Limited, Abuja, Nigeria for providing the fungal isolates used for this study.

