All published articles of this journal are available on ScienceDirect.

Vitality Structure of the Populations of Vegetative Motile Plants of Forest Ecosystems of the North-East of Ukraine
Abstract
Background:
The vitality level of the populations has turned out to be statistically reliably associated with such coenotic factors: age and density of forest stand. In general, the vitality spectra vary widely: the quality index Q of the populations ranges from 0,00 to 0,50, that is, it fully covers the theoretically possible scale of the values of this coefficient, which indicates the sensitivity of the vitality structure of the populations to the ecological-coenotic conditions and determines high informative value of the vitality analysis.
Objective:
The aim of the study is to assess the vitality structure of the populations of vegetative motile plant species – typical representatives of the grassy layer of forest ecosystems of the North-East of Ukraine as a factor that determines their stability and dynamics.
Methods:
The analysis of the vitality structure is based on the field studies of the populations of 4 vegetative motile species of plants – Aegopodium podagraria L., Asarum europaeum L., Carex pilosa Scop. and Stellaria holostea L. in forest ecosystems of the North-East of Ukraine. Vitality analysis was carried out according to Yu. A. Zlobin’s methodology. Vitality analysis procedure, classically, is carried out in three stages: 1) Selection of quantitative features that characterize the vital status of the individual plant; 2) Evaluation of vitality of individual plants that were included in the sample; 3) Integral assessment of the population’s vitality. Depending on the ratio in the population of plants of classes a, b and c, the population belongs to one of three vitality types: prosperous, equilibrium or depressive.
Results:
The obtained estimates of the vitality structure of populations of the clone-forming plants in the grass-shrub layer of forests of the North-East of Ukraine can be considered quite reliable, because they are based, in general, on a complete analysis of the morphological structure of about 13 thousand ramets of the studied species of plants. The statistical reliability of estimates of the population’s vitality structure is predominantly between 70 and 99% and only in some cases lower than 70%. As the clone grows older, its ramet’s vitality decreases and the clone degrades. New young clones, that start to form on the basis of genets, replace old ones. Such ramets have increased vitality, greater stress and competitive resistance. Due to the mechanisms of clone substitution in the living cover, the dominance of nemoralis herbs persists for a long time.
Conclusion:
The vitality spectra of the populations of the studied species of plants vary widely: the quality index Q of the populations ranges from 0,00 to 0,50, that is, it covers full theoretically possible scale of the values of this coefficient, which indicates the sensitivity of the vitality structure of the populations to the ecological-coenotic conditions and determines high informative value of vitality analysis. Prosperous populations: two populations A. europaeum of the associations Quercetum (roboris) coryloso (avellanae) – convallariosum (majalis) and Quercetum (roboris) coryloso (avellanae) – convallariosum (majalis), two populations A. podagraria of the associations – Pinetum (sylvestris) vacciniosum (myrtilli) and Querceto (roboris) – Pinetum (sylvestris) convallarioso (majalis) – vacciniosum (myrtilli), one population S. holostea of the association Querceto (roboris) – Pinetum (sylvestris) vacciniosum (myrtilli) and one population C. pilosa of the association Querceto (roboris) – Pinetum (sylvestris) vaccinioso (myrtilli) – convallariosum (majalis).
1. INTRODUCTION
Investigation of the state of the populations on the basis of population-structural approach is important in solving the problems of rational use, conservation, restoration and maintenance of the populations [1-7]. The study of the structure of plant populations provides extensive material that allows learning the adaptive properties of the species, obtaining the most complete picture of the mechanisms of stability and dynamics of plants of various ecological-coenotic groups [3, 8-13].
In a wide range of methods of population research the leading role is played by the study of the vitality structure of the coenopopulations of plants. High informativeness of analysis of the vitality state of populations is due to the fact that it relies not on one (size of the plant), but on a complex of morphometric parameters, which directly reflect growth and production process of plants [14-18].
In the comprehensive studies of vegetative motile species of grass plants in the forest ecosystems of the North-East of Ukraine, the issues of their systematics, evolution, ontogenesis [19-22] are widely covered, while the study of population characteristics is limited to a small list of regional works [23-25]. The study of the vitality structure of the populations was carried out only for some species. At the same time, determination of the mechanisms of stability of vegetative motile species, which are widely represented in the subshrub layer of forest ecosystems of the North-East of Ukraine and determine the first stages of the restoration of tree species, is an urgent and challenging task.
The aim of the study is to assess the vitality structure of the populations of vegetative motile plant species – typical representatives of the grassy layer of forest ecosystems of the North-East of Ukraine as a factor that determines their stability and dynamics. Plants of the highest class of vitality, having the highest viability, make in the population the largest contribution to both generative and vegetative reproduction. Vitality analysis of the populations of the studied plants will make it possible to identify the peculiarities of the change in the ecological-coenotic conditions of the habitat as it is the most sensitive and effective tool for such studies. In this way, such studies are part of a very important monitoring of the state of forest ecosystems for ensuring their proper protection and care, especially in the context of the bequest.
2. MATERIALS AND METHODS
The analysis of the vitality structure was based on the field studies of the populations of 4 vegetative motile species of plants – Aegopodium podagraria L., Asarum europaeum L., Carex pilosa Scop. and Stellaria holostea L. in forest ecosystems of the North-East of Ukraine. Syntaxons, for which typical are studied plant species, are given in Table 1.
Objects are typical of the forest region with domination of Quercus robur L. and addition of Pinus sylvestris L., Tilia cordata L., sometimes Acer platanoides, Betula pendula Roth. Such forests from the Querco-Tilieta nemoroherbosa group are close to the ecological phytocoenotic optimum for nemoralis herbs. This is their syntaxonomic or phytocoenotic optimum. We have selected phytocoenoses, in which dominant or codominant is one of the species of the studied nemoralis herbs: Aegopodium podagraria L., Asarum europaeum L., Carex pilosa Scop. or Stellaria holostea L. These species are herbaceous plants with deep roots that form clones.
Theoretical foundations and the algorithm of vitality analysis were formulated by Yu. A. Zlobin [18]. The procedure of vitality analysis, classically, is carried out in three stages. Detailed step-by-step technique was developed and presented by Yu. A. Zlobin [26]. At the first stage, in the population of plants in the individuals in the generative state, the maximum number of quantitative morphometric characteristics was taken into account. On these basis, allometric signs were calculated. Three or more signs were used to assess the vitality of individual plants, and then populations in general. The second stage was the assessment of the vitality of the individual plants that had been included in the sample. The procedure was carried out according to a strict mathematical algorithm – translation of the values of the signs, which was initially expressed in absolute values, in particles of a unit, definition of the vitality of the individuals. The vitality of individual plants lay in the amplitude from 0 to 1,0, and the higher it was, the higher was the vital state of this individual. In the main algorithm of the vitality analysis plants in the population were divided into three classes of vitality: “a”, “b” and “c”. For this, the range of values of plants’ vitality that lay in the amplitude from 0 to 1 was divided into three equal zones: more than 0,66 – the higher class of vitality – class “a”; the vitality of the individuals ranged from 0,33 to 0,66 – class “b” – intermediate; individual plants, whose vitality was less than 0,33 – class “c” – the lower class of vitality. In accordance with the values of the viability of the individual plants (Qindividual) they were referred to one or another class of vitality. The third stage was integral estimation of the populations’ vitality. Depending on the ratio in the population of individuals of classes a, b and c, the population belonged to one of the three vitality types: prosperous, equilibrium or depressive. In contrast to the vitality of individuals, the populations’ vitality lay in the amplitude of the values of Q from 0 to 0,5. This interval was divided into three equal parts: from 0 to 0,167, from 0,167 to 0,333 and from 0,333 to 0,500. The first of these intervals (Q less than 0,167) corresponded to depressive populations, the second (from 0,167 to 0,333) – equilibrium and the third (Q is more than 0,333) – prosperous.
The key attributes for establishing the vitality of individuals of the studied plant species were the following: total phytomass (W), Leaf Area (A), Reproductive Effort (RE).
In order to establish the vitality structure of populations Yu. A. Zlobin has developed a special software VITAL, which allows conducting step-by-step vitality analysis [16].
For all types of vegetative motile plants it is habitual to form clones, which are a set of ramets – the descendants of one genet. Within each clone, the placement of bushes of different age is not accidental, while in the ratio of partial shrubs of different age status clones differ significantly from each other.
Each ramet in the clones of the studied species of plants, in addition to evaluating its morphological structure, evaluated ontogenetic state in accordance with the generally accepted system of discrete description of ontogenesis. In order to evaluate the overall ontogenetic status of the clones, we have developed indices [25]: Index innovation – Iinn. (index of renewal), Index senilis – Isen. (aging index) and Index generative – Igen. (index of generativity). They were calculated by the following formulas:
1. Iinn. – ratio of pregenerative partial bushes to their total number:
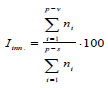 |
(1) |
where p, v, s – age states of partial bushes in standard notations.
In the index of renewal, the proportion of the pregenerative ramets of the clone indicates its active growth, which occurs by capturing a new territory.
2. Isen. – ratio of the partial bushes of the ontogenetic states g3, ss and s to their total number:
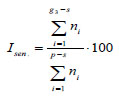 |
(2) |
3. Igen. – ratio of the number of young generative partial bushes (g1, g2) to their total number:
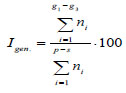 |
(3) |
The proportion of generative ramets of a clone is its generativity. It is higher where conditions of plant growth are most optimal, and ramets quickly transfer into the generative phase and retain the ability to flowering-fruiting annually for several years.
With the aim of integrative estimation of the age status of the populations of the studied species, Index aetas – Iaet. (index of the general age of populations) was developed and applied [25] in the form of the ratio of Index senilis to Index innovation of this population:
Iaet. = Isen. / Iinn.
The populations in the middle state of their age will have Iaet. equal to 1,0. Aging processes prevail when Iaet. is more than 1,0 and, accordingly, Iaet. less than 1,0 is characteristic for young populations.
| Associations |
|---|
| Asarum europaeum |
| I. Quercetum (roboris) coryloso (avellanae) – convallariosum (majalis) |
| II. Pinetum (sylvestris) asarosum (europaei) |
| III. Acereto (platanoiditis) – Quercetum (roboris) coryloso (avellanae) – aegopodiosum (podagrariae) |
| ІV. Quercetum (roboris) coryloso (avellanae) – convallariosum (majalis) |
| V. Querceto (roboris) – Pinetum (sylvestris) convallarioso (majalis) – vacciniosum (myrtilli) |
| VI. Pinetum (sylvestris) coryloso (avellanae) – vacciniosum (myrtilli) |
| Aegopodium podagraria |
| I. Quercetum (roboris) aegopodiosum (podagrariae) |
| II. Pinetum (sylvestris) vacciniosum (myrtilli) |
| III. Acereto (platanoiditis) – Quercetum (roboris) coryloso (avellanae) – aegopodiosum (podagrariae) |
| IV. Quercetum (roboris) aegopodiosum (podagrariae) |
| V. Querceto (roboris) – Pinetum (sylvestris) convallarioso (majalis) – vacciniosum (myrtilli) |
| VI. Pinetum (sylvestris) coryloso (avellanae) – vacciniosum (myrtilli) |
| Stellaria holostea |
| I. Acereto (platanoiditis) – Quercetum (roboris) caricoso (pilosae) – stellariosum (holosteae) |
| II. Betuletum (pendulae) stellariosum (holosteae) |
| II. Tilieto (cordatae) – Quercetum (roboris) stellariosum (holosteae) |
| IV. Querceto (roboris) – Pinetum (sylvestris) convallarioso (majalis) – vacciniosum (myrtilli) |
| V. Quercetum (roboris) coryloso (avellanae) – convallariosum (majalis) |
| VI. Querceto (roboris) – Pinetum (sylvestris) vacciniosum (myrtilli) |
| Carex pilosa |
| I. Betuletum (pendulae) caricosum (pilosae) |
| II. Quercetum (roboris) majanthemosum (bifolii) |
| III. Tilieto (cordatae) – Quercetum (roboris) stellariosum (holosteae) |
| IV. Quercetum (roboris)coryloso (avellanae) – convallariosum (majalis) |
| V. Quercetum (roboris) convallariosum (majalis) |
| VI. Querceto (roboris) – Pinetum (sylvestris) vaccinioso (myrtilli) –convallariosum (majalis) |
3. RESULTS
Asarum europaeum is a perennial grassy plant with a creeping rhizome that can reach a height of 5–10 cm. The rate of vegetative growth of Asarum europaeum is 2–5 cm per year. Stem is creeping, fluffy, short. Leaves are rounded, kidney-shaped. Flowers are single. Perianth is small. Fruit – a six-pin box. One generative plant produces up to 100 seeds a year. The plant is characterized by various forms of zoochory. Blossoms in April-May, seeds mature in June. Grows in shady broadleaf and mixed forests. Prefers fresh, moist, moderately humus, neutral or light-grained soils [25, 27]. Aegopodium podagraria – a plant with a horizontal creeping rhizome. The stem is straight, hollow. The upper leaves are almost naked, the lower – pubescent. Flowers are small, white. Inflorescence – a complex umbrella. Fruit – elongated flattened brown two-seeds, length 3-4 mm. One generative sprout gives from 2000 to 35 000 fruits. Blossoms from May to July, give fruit in July-August. It grows in deciduous and mixed forests, on felling, among shrubs, on fresh (in the lower layers of wet), slightly acidic (close to neutral), humus, rich, gravel, clay soils [25]. Stellaria holostea is a perennial grassy plant, 10 to 30 cm high, with a thin creeping branched rhizome. Stems are ascending. Leaves are opposite, narrow-banded. Flowers are situated on long painted stalks. Fruit – a spherical box, slightly shorter than a cup, one-snap, dry. Blossoms from mid-April to mid-June. Grows on humus, fresh, moist and well-ventilated forest soils, is often found in deciduous, coniferous and coniferous-deciduous forests of different types on fertile soils. Mesophyte [25]. Carex pilosa is a perennial grassy plant, 40–50 cm high, with long, underground creeping rhizomes. Two types of sprouts: shortened vegetative with broad (about 1 cm) and long two-component pubescent leaves and reproductive elongated, with five to seven short leaves (the length of their plates is 3–5 cm). Stems are triangular, smooth. The leaves are hibernating. Inflorescences have from 3 to 4 spaced spikes. Blossoms in the spring. Anemophile. Inflorescence consists of 3–4 spikes. The seeds ripen in July. Mesophyte. Mesotroph. Shadow-tolerant plant. The peak of photosynthesis falls in the spring, until autumn the intensity of photosynthesis gradually decreases. Gives fruit in April-May. It grows in deciduous (often broadleaf) and mixed forests. Prefers clay, loam or sandy, gray forest or rich sod-podzolic or black soils [25].
Asarum europaeum populations often dominate in the lower tiers of broadleaf and mixed forests in the North-East of Ukraine. In the forests of the North East of Ukraine, A. europaeum blooms in late April – early May. Aegopodium podagraria is one of the dominants of grass cover of deciduous forests. The plant does not belong to long-root species, but in the conditions of the north-eastern forests of Ukraine, it implements just this type of morphogenesis. Our studies and literature data [25] have shown that after the completion of active horizontal growth, such rhizomes begin to grow orthotropically and form overground partial shoots having rosettes of leaves. The result is a loose clone. The length of the long life cycle of A. podagraria reaches 50 years or more. In the absence of competitors, A. podagraria plants develop very rapidly. Rosette shoots with 9–12 leaves are formed already in the first year of life. In the nodes of the 5–8th leaves there are rhizomes up to 30 cm. The total length of rhizomes in a one-year plant reaches 2 m. At the end of the third year of life, one mother-plant forms up to 30 rosette partial bushes, some of which can already bloom in the third year. In A. podagraria, the root system consists of primary roots, which subsequently die quickly, and a significant number of adventitious. They are placed in the upper 25–30 cm of the soil horizons. Also, in plants A. podagraria there are thick fleshy roots, which carry out specific functions: supply of nutrients and retraction. Roots of A. podagraria are mycotrophic. The genet A. podagraria is represented by a system of monopodially growing rosettes (partial bushes) connected by plagiotropic rhizomes. Such rosettes are formed each year from the axillary buds of the shoots of previous orders. Lifetime of rosettes is one year. Stellaria holostea is a typical species for undisturbed oakeries and young aspen woods, is a typical representative of the European broadleaf flora. To the influence of ecological-coenotic factors various structural signs of S. holostea plants have different norm of reaction, as well as varied variations under different growth conditions. Therefore, it turns out that the system of correlation relations between morphometric parameters varies in different conditions in S. holostea. In more favorable conditions, the variability of the signs and correlation between them decreases. S. holostea is less demanding to the conditions of humidification, but more demanding to the fertility of the soil. The plant is resistant to trampling [25]. Carex pilosa is the most wide-spread dominant in the grass-bush-tree tier in mixed and broadleaved forests of the North-East of Ukraine. In the forest ecosystems of the North-East of Ukraine, flowering occurs in May.
The analysis of the vitality structure of the populations of A. europaeum was carried out on the basis of a sample of more than 3 thousand ramets. The results of the analysis of the vitality structure of A. europaeum populations are given in Table 2. It has turned out that from the studied A. europaeum populations, two of the associations of Quercetum (roboris) coryloso (avellanae) – convallariosum (majalis) and Quercetum (roboris) coryloso (avellanae) – convallariosum (majalis) were prosperous with a quality index Q equal to 0,35, and the other four are equilibrium with values of Q from 0,21 to 0,32. The statistical reliability of the obtained estimates was in the range from 50 to 99,5%.
| Associations | Quality Index Q |
Type of the Population | Level of Statistical Reliability, % | Iaet |
|---|---|---|---|---|
| I. Quercetum (roboris) coryloso (avellanae) – convallariosum (majalis) | 0,35 | Prosperous | 50,0 | 0,19 |
| II. Pinetum (sylvestris) asarosum (europaei) | 0,29 | Equilibrium | 99,5 | 0,14 |
| III. Acereto (platanoiditis) – Quercetum (roboris) coryloso (avellanae) – aegopodiosum (podagrariae) | 0,21 | Equilibrium | 70,0 | 2,35 |
| ІV. Quercetum (roboris) coryloso (avellanae) – convallariosum (majalis) | 0,35 | Prosperous | 80,0 | 0,60 |
| V. Querceto (roboris) – Pinetum (sylvestris) convallarioso (majalis) – vacciniosum (myrtilli) | 0,28 | Equilibrium | 70,0 | 0,65 |
| VI. Pinetum (sylvestris) coryloso (avellanae) – vacciniosum (myrtilli) | 0,32 | Equilibrium | 90,0 | 0,53 |
It should be noted that the population of A. europaeum of the lower vitality of Acereto (platanoiditis) – Quercetum (roboris) coryloso (avellanae) – aegopodiosum (podagrariae) was the oldest according to the ontogenetic composition and had the highest generative index of 68,5%.
In our opinion, the vitality structure of the populations of A. europaeum tends to decrease due to oppression by A. platanoides. This species has a more superficial root system, which leads to root competition between A. platanoides and A. europaeum. Another negative factor is the higher level of shading under the tent of the forest from Acer platanoides L. than in the pure forests of P. sylvestris and Quercus robur L.
For the analysis of the vitality structure of the A. podagraria populations, a sample of more than 2,000 ramets was used. In Table 3 the results of the analysis of the vitality structure of A. podagraria populations are presented. The statistical reliability of the data obtained is quite high and is not less than 70–90%.
The populations of A. podagraria from Pinetum (sylvestris) vacciniosum (myrtilli) and Querceto (roboris) – Pinetum (sylvestris) convallarioso (majalis) – vacciniosum (myrtilli) have appeared to be prosperous according to the vitality structure with a quality index Q of 0,41 and 0,39 respectively, and Acereto (platanoiditis) – Quercetum (roboris) coryloso (avellanae) – aegopodiosum (podagrariae) – depressive for Q, equal to 0,09. The last three populations were equilibrium with a quality index ranging from 0,20 to 0,26. In this species, in the studied populations, there was a decrease in the proportion of individuals of the higher class (A) and an increase in the proportion of individuals of the lower class (C). In our opinion, this is the result of the fact that in the North-East of Ukraine, the syntaxonomic optimum of A. podagraria falls on the light forests with the domination of Pinus sylvestris L.
The vitality structure of S. holostea populations is based on a sample of more than 2,000 ramets. The results of the analysis of the vitality structure of S. holostea populations are given in Table 4. The statistical reliability of the obtained estimates was in the range from 50 to 80%. Among the studied populations of S. holostea, one was prosperous with a quality index of 0,37, three equilibrium with a quality index from 0,23 to 0,27, and two – depressive with a quality index of 0,15–0,16. Depressive population of S. holostea from the association Betuletum (pendulae) stellariosum (holosteae) is the oldest according to the age composition with the highest value of the coefficient of age (Table 4). In this case, the connection of the increased age of the population with its depressive vitality structure was also recorded. At the same time, the depressive population from the association Quercetum (roboris) coryloso (avellanae) –convallariosum (majalis) is young with a coefficient of age of 0,62.
The analysis of the vitality structure of C. pilosa populations was carried out on the basis of a sample of about 3000 ramets. The results of the analysis of the vitality structure of C. pilosa populations are given in Table 5. The statistical reliability of the obtained estimates was in the range from 50 to 92,5%. The populations of C. pilosa in the sixth association were prosperous (quality index 0,35), four populations from the considered forest associations were equilibrium, with a quality index Q ranging from 0,24 to 0,28. Their vitality was practically the same. In contrast, C. pilosa population of the association of Betuletum (pendulae) caricosum (pilosae) was depressive with a Q index of 0,16. As in the case with S. holostea, it turned out that this population according to the ontogenetic composition of the individuals is the oldest Table 5.
| Associations | Quality Index Q |
Type of the Population | Level of Statistical Reliability, % | Iaet |
|---|---|---|---|---|
| I. Quercetum (roboris) aegopodiosum (podagrariae) | 0,20 | Equilibrium | 70,0 | 0,98 |
| II. Pinetum (sylvestris) vacciniosum (myrtilli) | 0,41 | Prosperous | 80,0 | 0,30 |
| III. Acereto (platanoiditis) – Quercetum (roboris) coryloso (avellanae) – aegopodiosum (podagrariae) | 0,09 | Depressive | 90,0 | 0,30 |
| IV. Quercetum (roboris) aegopodiosum (podagrariae) | 0,23 | Equilibrium | 70,0 | 0,30 |
| V. Querceto (roboris) – Pinetum (sylvestris) convallarioso (majalis) – vacciniosum (myrtilli) | 0,39 | Prosperous | 70,0 | 0,28 |
| VI. Pinetum (sylvestris) coryloso (avellanae) – vacciniosum (myrtilli) | 0,26 | Equilibrium | 90,0 | 0,36 |
| Associations | Quality Index Q |
Type of the Population | Level of Statistical Reliability, % | Iaet |
|---|---|---|---|---|
| I. Acereto (platanoiditis) – Quercetum (roboris) caricoso (pilosae) – stellariosum (holosteae) | 0,24 | Equilibrium | 80,0 | 0,73 |
| II. Betuletum (pendulae) stellariosum (holosteae) | 0,16 | Depressive | 50,0 | 3,56 |
| II. Tilieto (cordatae) – Quercetum (roboris) stellariosum (holosteae) | 0,23 | Equilibrium | 80,0 | 0,32 |
| IV. Querceto (roboris) – Pinetum (sylvestris) convallarioso (majalis) – vacciniosum (myrtilli) | 0,27 | Equilibrium | 80,0 | 0,63 |
| V. Quercetum (roboris) coryloso (avellanae) – convallariosum (majalis) | 0,15 | Depressive | 70,0 | 0,62 |
| VI. Querceto (roboris) – Pinetum (sylvestris) vacciniosum (myrtilli) | 0,37 | Prosperous | 70,0 | 1,82 |
| Associations | Quality Index Q |
Type of the Population | Level of Statistical Reliability, %% | Iaet |
|---|---|---|---|---|
| I. Betuletum (pendulae) caricosum (pilosae) | 0,16 | Depressive | 50,0 | 2,16 |
| II. Quercetum (roboris) majanthemosum (bifolii) | 0,25 | Equilibrium | 92,5 | 1,08 |
| III. Tilieto (cordatae) – Quercetum (roboris) stellariosum (holosteae) | 0,24 | Equilibrium | 80,0 | 1,13 |
| IV. Quercetum (roboris)coryloso (avellanae) – convallariosum (majalis) | 0,26 | Equilibrium | 99,5 | 0,79 |
| V. Quercetum (roboris) convallariosum (majalis) | 0,28 | Equilibrium | 70,0 | 0,94 |
| VI. Querceto (roboris) – Pinetum (sylvestris) vaccinioso (myrtilli) – convallariosum (majalis) |
0,35 | Prosperous | 80,0 | 1,0 |
For conducting vitality analysis, a group of species typical for forests region with domination of Q. robur, in which in herb-shrub tier dominate nemoralis species of herbs with predominance of vegetative motile clone-forming species, was taken. The results show that this tier can be viewed as a mosaic of clones of different ages and, accordingly, of different sizes, which dynamically substitute one another. Structural parts of the grass cover of nemoralis herbs in such cases are ramets of different ontogenetic and vital status, and separate genets that have not yet begun vegetative reproduction.
The obtained estimates of the vitality structure of the populations of clone-forming plants in the herb-shrub tier of forests of the North-East of Ukraine can be considered quite reliable, because they are based, in general, on a complete analysis of the morphological structure of about 13 thousand ramets of the studied species of plants. The statistical reliability of estimates of the populations’ vitality structure is predominantly between 70 and 99% and only in some cases lower than 70%. Such a few cases correspond to transitive types of populations’ vitality: prosperous-equilibrium or equilibrium-depressive.
As the clone is aging, ramets’ vitality decreases in it and the clone degrades. Such clones are replaced by new young ones that are just starting to form on the basis of genets. Such ramets have increased vitality, greater stress and competitive resistance. Due to mechanisms of clone substitution in the living cover, the dominance of nemoralis herbs is preserved for a long time: A. podagraria, etc. Thus, the mosaic of clones of nemoralis herbs is not static, but dynamic.
DISCUSSION AND CONCLUSION
Since vitality analysis of the population, as an integrity of ramets and their clones, is based on the vitality and morphological structure of individual plants, it accurately reflects the degree of compliance of the ecological and phytocoenotic conditions with bioecological characteristics of plants of this species. For example, in literature, it has been shown that in Convallaria majalis and Maianthemum bifolium, the first reaction to ecological and coenotic inhibition appears in deterioration of all morphological and growth parameters of plants, although the number of individuals in populations may even increase [28]. In the literature, there are many other data on the conditionality of the morphological status of plants under conditions of their growth. Summarizing the data and relying on the principle of allocation, M. H. Bashtovyi [29] argued that reducing the size of individuals and reducing metamers in them is reflection of adaptation of plants to stress conditions. Our studies and the results completely confirm this position.
Investigation of the vitality structure of populations of vegetative motile plants of the grassy layer in forest phytocoenoses has shown that more than half of the studied populations are equilibrium. The level of populations’ vitality proved to be statistically reliably associated with such coenototic factors as the age and density of the tree stand. In general, the vitality spectra vary widely: the quality index Q of the populations ranges from 0,00 to 0,50, that is, it covers full theoretically possible scale of the values of this coefficient, which indicates the sensitivity of the vitality structure of the populations to the ecological-coenotic conditions and determines high informative value of the vitality analysis. Prosperous populations are: two populations of A. europaeum in the associations of Quercetum (roboris) coryloso (avellanae) – convallariosum (majalis) and Quercetum (roboris) coryloso (avellanae) – convallariosum (majalis), two populations of A. podagraria in the associations – Pinetum (sylvestris) Vacciniosum (myrtilli) and Querceto (roboris) – Pinetum (sylvestris) convallarioso (majalis) – vacciniosum (myrtilli), one population of S. holostea from the association of Querceto (roboris) – Pinetum (sylvestris) vacciniosum (myrtilli) and one population C. pilosa from the association of Querceto (roboris) – Pinetum (sylvestris) vaccinioso (myrtilli) – convallariosum (majalis).
As a result of our research, we have shown that the vitality structure of the populations of a large group of vegetative motile plants is deteriorating with their distance from the ecological phytocoenotic optimum and is different in different types of forest phytocoenoses. In addition, it depends on the age of the ramets, which form clones.
ETHICS APPROVAL AND CONSENT TO PARTICIPATE
The reported experiments were in accordance with the Ethics Committee of Sumy National Agrarian University, Ukraine (No. 3445, dated 10-10-2019).
HUMAN AND ANIMAL RIGHTS
Not applicable.
CONSENT FOR PUBLICATION
Not applicable.
AVAILABILITY OF DATA AND MATERIALS
Not applicable.
FUNDING
None.
CONFLICT OF INTEREST
The authors declare no conflict of interest, financial or otherwise.
ACKNOWLEDGEMENTS
Declared none.

